999中国苗木网苗木供应信息:联系时请说明是在(999苗木网)上看到的苗木信息
东京野茉莉苗木的耐寒性研究 |
| 当前位置: 中国苗木网 > 栽培技术 > |
| 来源:未知 作者:admin 发布时间:2013-07-28 22:24 |
|
东京野茉莉(Styrax tonkinensis),又名越南安息香、白花树,属安息香科安息香属落叶乔木,主产越南东京湾,在我国主要分布于云南、贵州、广西、广东、湖南、福建和江西等地[1 ~ 3]。在原产地主要生长在海拔100 ~ l 000 m的低山丘陵地带,喜湿润疏松肥沃土壤,也耐贫瘠和耐干早[4 ~ 5]。该树种速生,具很强的萌芽更新能力,树干通直,木质白色细致,材轻软,木纤维较长,是理想的工艺、胶合板和纸浆用材;其种子含有人体所必需的油酸和亚油酸,具有较高的食用价值,树脂含香脂酸,为贵重药材,是一种很有发展前途的食药用油料树种[3 ~ 4];其花白色成串,清香,也是一种优良的绿化树种[6]。 999中国苗木网,999miaomu.com 1 材料与方法 1.1供试材料及其处理 供试材料为种源采自江西遂川,在江苏省镇江市扬子江林业科技有限公司苗圃生长的东京野茉莉一年生播种苗枝条。 将东京野茉莉枝条若干分别在-7℃、-18℃和-25℃下处理, 时间为 24、 48、 72 h。以5℃为对照, 处理结束后分别取枝条的韧皮部(避开芽眼)进行测定。 1.2试验方法 可溶性糖含量测定采用蒽酮比色法[9]。可溶性蛋白质含量测定采用考马斯亮蓝比色法[9]。脯氨酸含量测定采用酸性茚三酮比色法[9]。丙二醛(MDA)含量测定采用硫代巴比妥酸法[9]。POD活性测定采用比色法[9]。 2 结果与分析 2.1低温胁迫对东京野茉莉可溶性糖含量的影响 可溶性糖含量与植物抗寒力在多数植物中表现出正相关[10]。可溶性糖可以增加原生质浓度, 从而起到抗脱水作用及减少细胞内结冰机会,以此适应外界环境变化,减轻因低温而引起的冻害。 从图 1 可以看出在低温胁迫下,- 7℃处理 24 h 与72h后可溶性糖含量略有增加, 与对照差异不大。这是由于植株在5℃下时体内已聚集相当数量的糖分抵御低温。在-18℃和-25℃处理后可溶性糖含量下降,可见感受低温冻害后,植物可溶性糖含量有下降的趋势。 苗木网,www.999miaomu.com 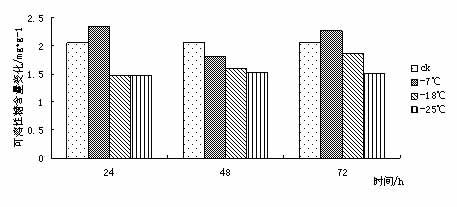 图1 低温胁迫对东京野茉莉可溶性糖含量的影响 2.2低温胁迫对东京野茉莉可溶性蛋白质含量的影响 植物在逆境下失水伤害是首要的[10]。可溶性蛋白质的增加可以束缚更多的水分,同时可以减少原生质内结冰而伤害致死的机会。 从图2可以看出在低温胁迫后,东京野茉莉幼苗韧皮部中可溶性蛋白呈先上升后下降的趋势,在-18℃处理24h、72小时后可溶性蛋白质含量达到最大值。可见可溶性蛋白含量的增加是东京野茉莉抗脱水、减少原生质内结冰的重要手段之一。 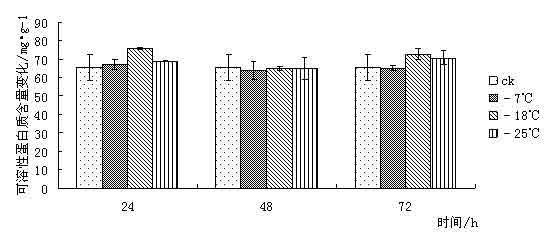 图二 低温胁迫对东京野茉莉可溶性蛋白质含量的影响 2.3低温胁迫对东京野茉莉POD活性的影响 正常情况下,植物体内存在着有效的保护酶系统(POD等),以清除活性氧等有毒物质。当植物体遇到低温胁迫时,这种清除机制就会受阻,导致体内活性氧的积累,造成对膜的伤害和对大分子的破坏。进而导致代谢功能的退化和细胞的死亡。 从图3可以看出,在低温胁迫后,东京野茉莉幼苗韧皮部POD活性均有所增加。在-25处理后的24h,48h,72h分别达到最大值,表现出东京野茉莉对低温有着较强的适应能力。 苗木网,999miaomu.com 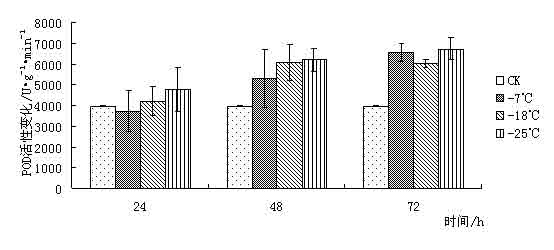 图3 低温胁迫对东京野茉莉POD活性的影响 2.4低温胁迫对东京野茉莉游离脯氨酸含量的影响 脯氨酸是植物体内一种重要的渗透调节物质,它的积累对适应逆境有重要意义,可以反映植物的受迫害状况。因此,其含量变化可以作为植物对逆境胁迫的一种生理生化指标[11]。 如图4所示,低温胁迫下,东京野茉莉幼苗韧皮部游离脯氨酸含量明显增加。随着温度的不断降低, 游离脯氨酸含量有逐渐增加后降低的趋势并在-18℃处理48后达到最大值。可见东京野茉莉通过增加游离脯氨酸绝对含量来提高其抗寒性。 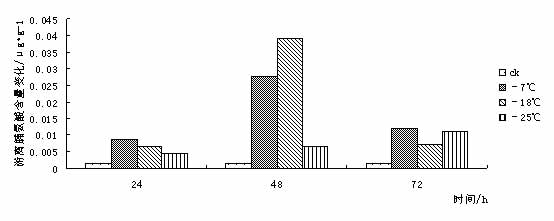 图4低温胁迫对东京野茉莉游离脯氨酸含量的影响 2.5低温胁迫对东京野茉莉MDA含量的影响 丙二醛 (MDA)是生物膜系统膜脂过氧化产物之一,其含量不仅表示过氧化程度和膜系统的伤害程度,而且也可以间接地表示组织中自由基的含量[ 12 ]。 从图5可以看出,在24h处理下,随着温度的下降MDA含量出现先升高后降低的趋势,说明低温胁迫下,脂质发生过氧化作用,细胞膜受损。在48h,72h处理下,低温胁迫后的东京野茉莉幼苗韧皮部MDA含量均有所下降,这说明随着胁迫时间的正常,东京野茉莉逐渐修复细胞膜,以达到抗寒的目的。 中国苗木网,999miaomu.com 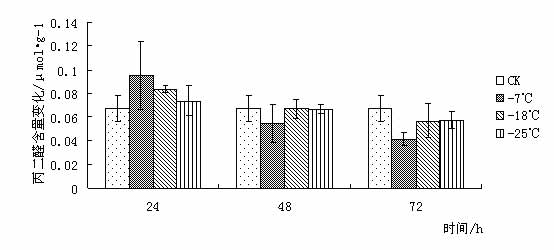 图5低温胁迫对东京野茉莉MDA含量的影响 3 讨论 低温冰冻会引起一系列细胞代谢功能的改变,人们采用可溶性糖含量、可溶性蛋白质含量、脯氨酸含量、SOD活性、POD活性、丙二醛含量、NR活性等研究林木抗寒性[13]。 3.1可溶性糖含量与东京野茉莉的抗寒性 研究结果表明可溶性糖可以增加原生质浓度, 从而起到抗脱水作用及减少细胞内结冰机会,对原生质,冻敏感蛋白质、偶联因子等起到保护作用[10]。在发生低温时,大部分植物都会积累一定量的可溶性糖来适应低温带来的逆境。本实验结果表明在-7℃低温处理下东京野茉莉可溶性糖含量有所增加,说明在-7℃时东京野茉莉可通过升高其可溶性糖含量来增加原生质浓度从而抵御低温胁迫。 3.2可溶性蛋白质含量与东京野茉莉的抗寒性 研究结果表明可溶性蛋白质的增加可以束缚更多的水分,同时可以减少原生质内结冰而伤害致死的机会[10]。本试验结果表明在低温胁迫时,东京野茉莉通过可溶性蛋白的积累,从而降低水势,增强渗透调控能力。 3.3 POD活性与东京野茉莉的抗寒性 正常情况下,植物体内存在着有效的保护酶系统,以清除活性氧等有毒物质。当植物体遇到低温胁迫时,这种清除机制就会受到阻碍,导致体内活性氧的积累,造成对膜的伤害和大分子的破坏,使DNA产生损伤,影响蛋白质的合成与稳定,进而造成代谢功能消失时和细胞死亡,一般认为,低温胁迫下POD活性表现为先升后降[14]. 苗木网,999miaomu.com 本试验中,在低温胁迫下东京野茉莉POD活性呈先上升后下降趋势,且随着时间的增长有较大幅度的升高,表明东京野茉莉在低温胁迫下保护酶可以维持较高水平,从而避免了活性氧等自由基的大量积累,减轻了植物体内活性氧带来的伤害。 3.4游离脯氨酸含量与东京野茉莉的抗寒性 研究结果表明,脯氨酸在植物抗冻中具有重要作用,随着温度的不断降低,植物叶片中脯氨酸含量呈明显增加趋势。越抗寒的品种增加的倍数越高。[11].脯氨酸是植物体内水溶性最大的氨基酸之一,在低温胁迫下,游离脯氨酸的积累在适应逆境中起到一定作用。本试验中,在低温胁迫下游离脯氨酸含量呈先上升后下降趋势,在-18℃处理24h的组分中达到最大值,说明游离脯氨酸的增加时东京野茉莉抗寒反应的一项重要手段。 3.5 MDA含量与东京野茉莉的抗寒性 丙二醛 (MDA)是生物膜系统膜脂过氧化产物之一,其含量不仅表示过氧化程度和膜系统的伤害程度,而且也可以间接地表示组织中自由基的含量。刘俊英等在研究低温胁迫对雪松膜脂过氧化物及保护酶的影响时,证实了植物体内 MDA含量可以反映植物膜伤害及衰老程度[11].本试验中,低温胁迫下MDA含量随着时间的变化呈现先升高后降低的趋势,这表明在低温下东京野茉莉细胞膜受到一定程度的破坏,后在POD等酶的作用下逐渐使活性氧代谢的平衡恢复。说明机体抗氧化系统的调节能力较强。 苗木网,999miaomu.com 在可溶性糖、可溶性蛋白含量增加和脯氨酸、POD等保护酶的共同作用下,东方东京野茉莉就能对低温胁迫产生较强的抵抗能力, 但我们同时需指出上述所使用的 5个指标中,只有游离脯氨酸具有极显著差异,其他 4个指标均差异不显著。说明在该研究中游离脯氨酸是测定植物抗寒性的重要指标。应该指出,具有抗寒力的植物一定存在着抗寒基因[15], 在低温下表达出生理和形态结构的抗性。生理抗性也不是单一因子的作用,可能是综合的生理因素[16]。因此上述测试的几种生理因子与东京野茉莉抗寒性有一定关系,是否与其它方面有关系还有待进一步探讨。 在该研究中,有许多尚不能确定的认识,反应了植物抗寒性机制的复杂性,需要进行深入研究。植物生活在光、温、 水、 气、 热共同作用的环境中,低温胁迫只是其中研究的一个方面,因此在加强单一因子研究的基础上,研究双因子或多因子共同作用,才能达到引种成功和适地适树的目标。 参考文献 [1] 中国科学院植物研究所. 中国高等植物图鉴第三册[M]. 北京:科学出版社,1985. 338. [2] 郑万钧. 中国树木志第二卷[M]. 北京:中国林业出版社,1985. 12. 1 602-1604. 999苗木网,999miaomu.com [3] 傅立国,陈潭清.中国高等植物第六卷[M]. 青岛出版社,2003,10. 29. [4] 肖复明,曾志光,杨桦,等. 东京野茉莉种子油营养成分研究[J]. 天然产物研究与开发,2005,17(3):344-345. [5] 吴克选,曾志光,周小平,等. 东京野茉莉野外调查报告[J]. 江西林业科技,2002(2):25-27. [6] 戴晓龙. 东京野茉莉及繁育技术[J]. 特种经济动植物,2002(6):21. [7]严青,马玉涛,施建军,等.低温胁迫对3 种牧草幼苗抗性生理指标的影响[J ].青海大学学报,2007 ,25(1) :54-57. [8]黎明,李福秀,马焕成,等.香木莲对短时低温胁迫处理的生理生态响应[J ].北方园艺,2006(1) :37-39. [9]张治安、陈展宇,植物生理学实验技术,吉林大学出版社,2008.7 100,110,182,192. [10]刘祖祺等.植物抗性生理学.北京:中国农业出版社,199 [11]闫中园, 金研铭.不同品种紫斑牡丹的抗寒性研究与比较[J ].安徽农业科学,2009, 37 (24) :11511 - 11513. [12]刘俊英,姚科云,冯耀飞,等.低温胁迫对雪松膜脂过氧化及保护酶的影响[ J ] .山西农业大学学报, 2004 (4) : 399 - 400 . 999中国苗木网,www.999miaomu.com [13]王利,丰震,张东宁.国内林木抗寒性研究的进展及展望[J].山东林业科技,2002(1):48-49. [14] 谭大海,罗新义,沙伟.低温胁迫下4个苜蓿引进品种 SOD、 POD活性的变化[J ].中国饲料,2007(1) :25-31. [15]9. Nordin et. al. Separate signal pathways regulate the expression of a low-temperature- induced gene in Arabidopsis thaliana(L.) Heynh, Plant Molecular Biology, 1991,(16): 1061-1071 [16]万清林等.侧金盏花成株年生长节律生理特性的研究.植物研究,1996,(3):351-355. 苗木网,999miaomu.com 作者:王航 肖雅 导师:喻方圆 999苗木网,999miaomu.com |
| 上一篇:鲜切花选购要诀 下一篇:盆栽缠线的要领须知 |